|
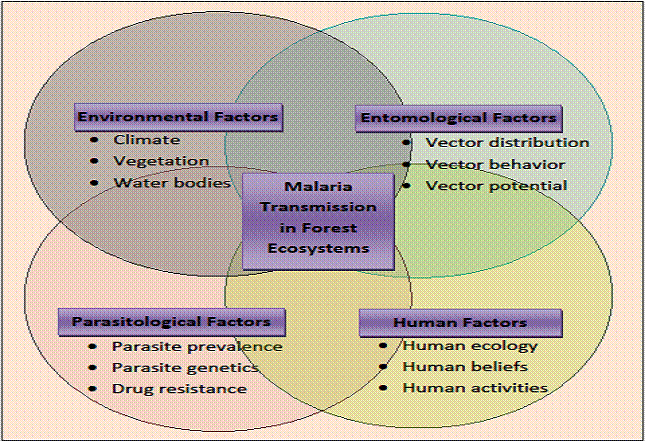
Tổng quan về động lực lan truyền sốt rét ở các hệ sinh thái rừng trên thế giới
Mặc dù trong vài thập niên qua, thế giới đã đạt được nhiều thành trong việc kiểm soát bệnh sốt rét, giảm cả số mắc và số tử vong đáng kể, cơ cấu ký sinh trùng có xu hướng đảo ngược ở một số quốc gia và một số vùng trên thế giới. Song, hiện nay, bệnh sốt rét vẫn còn là mối quan tâm đối với sức khỏe cộng đồng do phải đối mặt với nhiều thách thức mới như muỗi kháng với hóa chát diệt, ký sinh trùng kháng với thuốc ưu tiên trong điều trị, sốt rét biên giới, sốt rét ở nhóm dân di biến động là các hình thái sốt rét mà toàn cầu đang phải tìm cahcs nghiên cứu khắc phục, giảm thấp, tiến tới loại trừ sốt rét, đặc biệt là sốt rét do P. falciaprum. Mô hình lan truyền bệnh sốt rét đã có nhiều thay đổi, do vậy các chiến lược phòng chống cũng đã có một số thay đổi theo cho phù hợp. Để có một cái nhìn tổng quan về lan truyền sốt rét trên toàn cầu, chúng tôi giới thiệu về động lực lan truyền bệnh sốt rét ở các hệ sinh thái rừng trên thế giới của nhóm các tác giảNarayani Prasad Kar,Ashwani Kumar,Om P Singh,Jane M Carltonvà Nutan Nanda đăng trên tạp chíParasites & Vectors (2014). Sự phân tầng sốt rét toàn cầu Sốt rét là bệnh truyền nhiễm cấp tính do ký sinh trùng sốt rét thuộc giống Plasmodium. Hiện nay, bệnh lưu hành tại 104 quốc gia vùng nhiệt đới và cận nhiệt đới với hơn một nửa dân số thế giới (3,4 tỷ người), trong đó có khoảng 2,57 tỷ người có nguy cơ nhiễm KSTSR P. falciparum [2], và 2,5 tỷ có nguy cơ nhiễm P. vivax [3]. P. malariae và P. ovale hai loài KSTSR đóng góp một phần rất nhỏ trong tổng số ca mắc bệnh nhưng dân số có nguy cơ nhiễm P. malariae tập trung chủ yếu ở khu vực cận Sahara châu Phi, khu vực Đông Nam Á, các đảo tây Thái Bình Dương, và Amazon Basin [4, 5 ]. P. ovale là phổ biến ở châu Phi [5] và nó cũng được báo cáo từ khu vực châu Á-Thái Bình Dương [6]. P. knowlesi là loài ký sinh trùng thứ năm [7] vốn trước đây thường nhiễm ở động vật linh trưởng nhưng hiện nay đã có báo cáo nhiễm ở người từ các vùng rừng núi xa xôi của các quốc gia Đông Nam Á [8-11] như Malaysia, Indonesia, Papua New Guinea, Việt Nam,…. Sốt rét rừng (Forest malaria) Định nghĩa "rừng" “Kiểu sinh thái rừng - forest ecotype” được UNESCO định nghĩa như sau: “Kiểu sinh thái rừng” có địa hình với độ bao phủ tán cây hơn 10% và diện tích hơn 0,5 ha, trong đó rừng tự nhiên và rừng trồng [12] với chiều cao cây tối thiểu là 5 m, bao gồm cây cà phê, cao su, nút chai gỗ sồi, cây ăn quả và trồng rừng chắn gió và vanh đai bảo vệ hơn 20 m chiều rộng [13]. Thảm thực vật rừng được phân loại gồm rừng mưa, rừng rụng lá, rừng cây bụi, rừng mưa cao nguyên, rừng núi vùng cao [14]. Rừng mưa, rừng rụng lá, rừng cây bụi thường phân bố ở vĩ độ thấp đến trung bình và rừng mưa cao nguyên, rừng núi vùng cao là một phần của quần xã sinh vật trên cao. Tác động của bệnh sốt rét rừng Các hệ sinh thái rừng có vai trò rất lớn trong việc hỗ trợ lan truyền bệnh sốt rét, góp phần đáng kể vào gánh nặng bệnh tật trên phạm vi toàn cầu. Một đánh giá toàn cầu cho biết "các rừng kín trong các khu vực nguy cơ sốt rét bao phủ khoảng 4,8 triệu km2" [12]. Ước tính có khoảng một nửa dân số nguy cơ mắc bệnh sốt rét xảy ra ở những người sống ở các khu vực rừng (1,4 tỷ người) chiếm 11,7, 18,7, 35,1 và 70,1 triệu người tương ứng từ 1,5 triệu km2 ở vùng Amazon, 1,4 triệu km2 ở Trung Phi, 1,2 triệu km2 ở Tây Thái Bình Dương, và 0,7 triệu km2 trong khu vực Đông Nam Á [15,16]. Tương ứng với các khu vực rừng chứa đựng những khu nguy cơ sốt rét là 11.16 - 15.71 triệu km2, 6.53 - 7,80 triệu km2, 1,93 - 5.19 km2, 2,70 - 2,72 triệu km2 [12,15,16]. Kiểm soát bệnh sốt rét ở các khu rừng trên thế giới là một thách thức lớn [17] mà hiện nay các nghiên cứu dù đã tập trung và đầu tư tiền cửa và nhân lực, song vẫn chưa đưa ra một giải pháp nào tổng thể và hiệu quả trên phạm vi diện rộng trên khu vực hoặc toàn cầu. Những nguy cơ tiềm ẩn sốt rét rừng Hầu hết các nghiên cứu liên quan đến sốt rét rừng thường được tập trung vào các yếu tố địa phương gắn liền với lan truyền bệnh sốt rét như khoảng cách từ nhà đến rừng, tác động của việc phá rừng và tái trồng rừng, ảnh hưởng của rừng đến vi khí hậu, sinh thái học véc tơ, khả năng tồn loài Plasmodium spp. và các hoạt động của con người trong các khu rừng như thế nào. Trong bài này, sẽ phân tích các yếu tố cơ bản ảnh hưởng đến lan truyền bệnh sốt rét ở các khu rừng trên toàn thế giới. Muỗi phân bố khác nhau tùy theo vị trí rừng và chúng có thể thay đổi tập tính, hành vi phù hợp với môi trường và điều kiện vi khí hậu [18], dân số và tập quán cộng đồng [19-20]. Những cộng đồng sống gần rừng thường là các bộ tộc, nơi có cơ sở hạ tầng giao thông, y tế nghèo nàn. Một số hoạt động của con người như nương rẫy, ngủ trong rừng để khai thác lâm sản rừng, săn bắn, xây dựng nhà và chăn nuôi gia súc, làm gia tăng nguy cơ nhiễm bệnh sốt rét. Đây là thách thức lớn để giáo dục cộng đồng về phòng chống sốt rét rừng và nếu không có sự hợp tác của cộng đồng thì rất khó để kiểm soát bệnh sốt rét [21]. Giờ đây, các cộng đồng có nguy cơ mắc sốt rét trên toàn thế giới đang tập trung các nguồn lực vào việc loại bỏ/ tiêu diệt sốt rét [22, 23]. Tuy nhiên, chúng ta muốn thành công thì cần ngăn ngừa sự xuất hiện trở lại hoặc tái xuất hiện các ổ dịch tiềm ẩn/ không kiểm soát bệnh sốt rét rừng [22],[24]. Bệnh sốt rét đã giảm đáng kể trong thời kỳ xóa bỏ bệnh sốt rét trước đây tại nhiều khu vực trên thế giới, nhưng sau đó bệnh phục hồi và gây ra hậu quả nặng nề đối với sức khỏe cộng đồng [23],[25],[26],[27]. Vấn đề liên quan đến bệnh sốt rét rừng rất phức tạp, do các ổ chứa nhiễm sốt rét tìm ẩn mà không được giải quyết một cách triệt để [28],[29]. Nguồn gốc và sự phát triển kháng thuốc sốt rét và kháng hóa chất diệt côn trùng thường được tìm thấy có liên quan đến rừng và các khu vực gần rừng [24],[30],[32]. Ngoài ra, sự có mặt các trường hợp nhiễm bệnh không có biểu hiện lâm sàng, sự hiện diện ký sinh trùng sốt rét ở động vật linh trưởng và véc tơ truyền ký sinh trùng sốt rét từ động vật sang người có thể đặt ra những thách thức bổ sung đối với sức khỏe con người tại các khu vực rừng và bìa rừng [33],[35], đây là những nơi giám sát bệnh sốt rét thường hạn chế [33],[36]. Thỉnh thoảng các đợt bùng phát dịch sốt rét (lan truyền bệnh không ổn định) có thể xảy ra khi lan truyền bệnh sốt rét mở rộng từ rừng núi (ổ bệnh) đến các vùng ven đô và đô thị [37], nơi mật độ dân số đông và sự hiện diện các véc tơ có thể góp phần gây là dịch bệnh lớn. Những yếu tố chính làm cho sinh thái rừng khác so với các hệ sinh thái khác Các yếu tố chính phân biệt hệ sinh thái rừng với các hệ sinh thái khác liên quan đến động lực truyền bệnh sốt rét, đó là những ảnh hưởng của rừng đối với nhiệt độ, lượng mưa [38], độ ẩm [39-41], độ che phủ [42], hệ thực vật, động vật [43], hàm lượng chất hữu cơ trong các vùng nước muỗi đẻ [44] và thiếu cơ sở hạ tầng [45]. Trong các khu vực rừng có nhiều khó khăn để phát triển cơ sở hạ tầng do địa hình không đồng nhất, sự có mặt nhiều dòng suối và thảm thực vật dày đặc. Ngoài ra, giao tiếp kém gây cản trở cho hoạt động phòng chống sốt rét đặc biệt là vào mùa mưa [21,45]. Hơn nữa, khu rừng với nhiều đồi núi là điều kiện thuận lợi cho lan truyền bệnh sốt rét hơn do các sườn nùi tạo thành những dòng suối nhỏ tạo điều kiện thuận lợi cho véc tơ sốt rét sinh sản và phát triển hiệu quả hơn [46]. Rừng ảnh hưởng đến sinh thái học và phân bố véc tơ, cũng như sự phân bố của các loài ký sinh trùng sốt rét. Khu vực rừng chủ yếu là nơi sinh sống của các bộ tộc [47,48], họ thường mù chữ và chỉ tin vào các truyền thống và tập quán cũ kỹ, lạc hậu và họ thường sợ hãi thế giới bên ngoài, điều này dẫn đến sự phụ thuộc vào cách điều trị bệnh sốt rét của người bản địa [49]. Ảnh hưởng các thông số địa hình trong các khu vực rừng Các yếu tố địa hình ảnh hưởng tới động lực lan truyền bệnh sốt rét ở các khu rừng khác so với các vùng khác [38,40,50-52] vì rừng là môi trường ẩn nấu, véc tơ sốt rét thích nghi [53,54] điều này phản ứng khác nhau đối với những thông số do cấu trúc gen của chúng và sự hiện diện rừng ảnh hưởng lớn hơn so với khu vực ít hoặc không có rừng [18, 28, 52, 55]. Nhiệt độ, lượng mưa và ẩm độ Ở vùng rừng cao nguyên Đông Phi, nhiệt độ trung bình trong tháng trước và lượng mưa trong hai tháng trước đó đã cho thấy một mối quan hệ tuyến tính bậc hai với mật độ Anopheles gambiae [51]. Một nghiên cứu khác trong cùng khu vực cho thấy rằng tỷ lệ giữa lượng mưa và lượng thoát hơi tiềm năng là động lực cho gia tăng mật độ An. gambiae và An. arabiensis [40]. Cũng giống như nghiên cứu véc tơ ở Gambia đã cho thấy sự gia tăng mật độ nhanh vào cuối mùa khô và tối đa sau khi bắt đầu những cơn mưa làm cho độ ẩm tăng lên [56]. Nhìn chung cây cối ở trong rừng cộng thêm hơi ẩm trong không khí do sự bốc hơi nước và giúp giảm nhiệt độ, do đó làm tăng lượng mưa. Môi trường ẩm và các vị trí sinh sản của muỗi được tạo ra do mưa làm gia tăng mật độ, tuổi thọ của véc tơ và do đó làm tăng lan truyền bệnh sốt rét [19]. Thảm thực vật Thảm thực vật gần nơi ở con người làm gia tăng quần thể véc tơ sốt rét rừng và do đó làm tăng lây truyền bệnh sốt rét [57],[58],[59]. Những Làng có rừng lá rộng hơn và thảm thực vật đất ngập nước ở Belize và các làng trong rừng của Bangladesh có tỷ lệ mắc bệnh sốt rét cao hơn [60,61] do mật độ véc tơ trong rừng cao [61]. Các véc tơ rừng thường thích tán cây che phủ [42,62] và các hốc cây là nơi trú ẩn của các véc tơ sốt rét [63],[64]. Hệ thực vật rừng và cây lấy đường cũng đã được chứng minh là yếu tố quan trọng quyết định đến năng lực véc tơ. Sự sẵn có thực vật lấy đường làm gia tăng số trứng [43,65] và tăng khả năng tồn tại của loài An. gambiae là nguyên nhân làm gia tăng lan truyền bệnh sốt rét [66]. Ngoài ra, lá cây rơi vào môi trường sống của bọ gậy tạo ra môi trường vi khí hậu ổn định và thức ăn cho bọ gậy, trong đó phố biến là véc tơ An. dirus ở Đông Nam Á [52]. Nguồn nước Muỗi trưởng thành trong các nguồn nước (môi trường sống của bọ gậy/ lăng quăng) và phân tán theo khoảng cách bay của chúng. Ví dụ, quần thể muỗi An. gambiae và An. funestus được quan sát thấy giảm khi gia tăng khoảng cách từ sông Yala ở quốc gia Kenya [42]. Ngay cả một thay đổi nhỏ trong khoảng cách từ các nguồn nước có thể ảnh hưởng đến lan truyền sốt rét [19, 67]. Anopheles fluviatilis [68], Anopheles maculatus [68,69] và An. minimus [52, 68, 69] phổ biến gần dòng suối trong khu vực rừng có khí hậu mát và tán cây [70], nhưng bọ gậy An. dirus phát triển tốt trong những vũng nước nhỏ, sạch và nguồn nước ứ đọng trong các vùng rừng châu Á [52, 68]. Tại châu Phi, bọ gậy An. gambiae s.sphát triển tốt hơn ở các nguồn nước dưới tán rừng rậm rạp hơn là dưới tán rừng thưa [71]. Nói chung, bọ gậy của véc tơ rừng phát triển tốt hơn trong các nguồn nước dưới tán cây, nơi nhiệt độ nước ổn định và thường dao động từ 3 - 3,50C thấp hơn so với các nguồn nước tiếp xúc với ánh nắng mặt trời [71]. Phá rừng Khi phá rừng con người làm giảm độ che phủ của tán rừng do đó làm tăng tiếp xúc ánh sáng mặt trời với các nơi sinh sản, nghỉ ngơi của véc tơ, do đó làm thay đổi môi trường sống của véc tơ. Các nghiên cứu cho thấy, các loài muỗi An. dirus [52,72], An. fluviatilis [72,73], An. minimus [72,74], và An. funestus [74,75], An. darlingi [72] ưa thích môi trường sống có nhiều tán cây rừng và ngược lại một số loài như An. gambiae [72,75], và An. maculatus [68,69,76] ưa thích môi trường sống có ánh nắng mặt trời. Thay đổi mật độ Anophelines do nạn phá rừng đã được báo cáo trên toàn thế giới và nó liên quan đến môi trường sống rộng và ưa thích ánh sáng mặt trời đã được xem xét trong phương pháp phân tích tổng hợp/ lập bảng [72] và đã được tìm thấy những thay đổi mật độ Anophelines và tỷ lệ mắc bệnh sốt rét thay đổi theo loại hình phát triển, nông nghiệp và địa phương[72]. Điều này đã được dự báo rằng nạn phá rừng ở miền trung châu Phi và vùng nhiệt đới châu Mỹ có thể làm tăng bệnh sốt rét [12],[77], trong khi ở châu Á thì một số nơi cho thấy nạn phá rừng sẽ dẫn đến giảm sốt rét [78]. Đúng như dự báo ở khu vực Sahara, tỷ lệ mắc sốt rét tăng do nạn phá rừng như một hệ quả của việc gia tăng mật độ véc tơ An. gambiae và An. arabiensis [72] và sự gia tăng quần thể An. funestus và An. gambiae ở cận Saharan Châu Phi [72]. Tương tự như vậy phá rừng làm gia tăng quần thể véc tơ An. darlingi và An. aquasalis [72] ở Nam Mỹ, đi kèm với sự gia tăng bệnh sốt rét ở Guyana và Amazonia [72]. Việc dự báo bệnh sốt rét giảm ở các khu vực phá rừng châu Á có thể là do sự sụt giảm (halophobic) các véc tơ rừng như An. dirus ở Thái Lan và An. fluviatilis ở Ấn Độ [73,79]. Tuy nhiên, lan truyền bệnh sốt rét đã được đẩy mạnh bởi loài An. minimus do nạn phá rừng ở Thái Lan và Ấn Độ[80], cũng như An. culicifacies ở Nepal và Sri Lanka [72] và An. philippinensis, An. annularis và An. Varuna ở Ấn Độ [80]. Nguy cơ sốt rét gia tăng liên quan đến sự tồn tại tình trạng phá rừng nếu như véc tơ An. darlingi có mặt trong môi trường sống phát quang [81], khi tỷ lệ đốt của An. darlingi được ước tính gia tăng 278 lần ở khu vực rừng bị tàn phá [82]. Do đó nạn phá rừng ảnh hưởng đến lan truyền sốt rét phụ thuộc vào sự đa dạng của các véc tơ sốt rét ở một khu vực nhất định. Các chỉ số về côn trùng trong khu vực có rừng Tác động của rừng và trồng rừng đến sự phong phú của véc tơ Các véc tơ trong vùng sinh thái rừng phản ánh sở thích hay khả năng thích ứng của véc tơ với kiểu sinh thái rừng. Động lực lan truyền bệnh sốt rét rừng có thể là kết quả của sự phong phú véc tơ [83]. Các loài véc tơ khác nhau hoặc các loài “anh em” cùng khu vực phân bố có thể có mặt trong một khu vực cụ thể mà các quần thể biến động theo mùa [52],[84]. Trong rừng rậm, đồi núi khu vực biên giới Thái Lan - Myanmar, loài muỗi An. dirus đã được tìm thấy vào đầu mùa mưa trong khi nhiều loài An. baimaii đã được tìm thấy ở đoạn giữa mùa mưa [52]. Tuy nhiên, phần phía Nam Thái Lan ít có rừng, loài An. cracens phổ biến hơn An. baimaii vào đầu mùa mưa mặc dù vào cuối mùa mưa An. scanloni phổ biến hơn loài An. cracens [52,84]. Ở vùng rừng núi cận nhiệt đới, phía tây bắc Argentina, loài An. argyritarsis phong phú hơn loài An. pseudopunctipennis và cả hai véc tơ đạt mức cao nhất trong mùa xuân [85]. Trong số các loài thuộc phức hợp loài gambiae, loài An. gambiae s.s. ít thích nghi với điều kiện nóng hơn so với loài An. arabiensis [86],[87], vì thế trước đây loài này phong phú trong rừng hơn so với sa mạc, theo phản ánh phân bố không gian và thời gian của véc tơ ở châu Phi [29],[54],[88],[90]. Trong mùa khô, khi quần thể véc tơ rừng chính (ví dụ, An.gambiae ss) giảm, các véc tơ phụ (ví dụ: An. arabiensis) đóng vai trò lan truyền bệnh sốt rét. Rừng nhân tạo bao gồm cả những khu vực rừng trồng lớn đáng kể hoặc rừng tái sinh cũng gây ra sự thay đổi môi trường sống và ảnh hưởng đến sự phong phú véc tơ sốt rét dẫn đến thay đổi trong kịch bản lan truyền bệnh sốt rét. Ví dụ sốt rét tăng do đồn điền cà phê ở Thái Lan [91], các đồn điền cọ ở Cameroon [92], Papua New Guinea [93] và Malaysia [94]; đồn điền cao su tại Cameroon [95], Thái Lan [91] và khu vực trồng cây ăn quả ở biên giới Thái - Myanmar và các khu vực Đông Nam Á khác [91,96,97]. Rừng trồng thương mại và tái trồng rừng, làm gia tăng hoạt động của con người điều này làm gia tăng tiếp xúc giữa con người với véc tơ truyền bệnh sốt rét ở những khu vực này [78],[91],[96],[97]. Tập tính các véc tơ thích nghi với rừng Một số véc tơ không có rừng khác biệt với các véc tơ rừng điều này thể hiện sự thay đổi trong mô hình dãi băng nhiễm sắc thể và cho thấy sự thay đổi sinh thái học so với các hình thức véc tơ không có rừng [53],[54],[98]. Sự thay đổi các hình thức véc tơ đi kèm với sự khác biệt khả năng véc tơ, thói quen đốt mồi và khác biệt kháng với hóa chất diệt côn trùng, do đó ảnh hưởng đến chiến lược phòng chống véc tơ [99],[100] đáp ứng với lan truyền bệnh sốt rét [98],[101]. Obsomer và cộng sự (2007), trong bài đánh giá về loài muỗi An. dirus ở khu rừng châu Á, tác giả nhấn mạnh môi trường rừng, hành vi con người và việc sử dụng hóa chất diệt côn trùng đối với véc tơ di truyền để giải thích cho hành vi không đồng nhất của véc tơ [52], mà còn hỗ trợ các quan điểm trước đó của Trung và cộng sự [18],[28]. Họ xem xét sự không đồng nhất về hành vi muỗi Anophelines ở Đông Nam Á liên quan đến rừng, đồi núi và các yếu tố khác và thấy rằng sự thay đổi hoạt động đốt người vào đầu buổi tối của An. dirus A (An dirus.) và An. minimus A (An. minimus Theobald) và mức độ hoạt động ngoài nhà cao hơn tỷ lệ nghịch với khoảng cách từ các khu rừng và đồi [52]. Hình thức An. gambiae rừng cho thấy thích sống ngoài nhà hơn ở miền nam Sierra Leone trong khi các hình thức Savannah hầu hết thích sống trong nhà [98]. An. dirus đốt máu ban ngày cũng được quan sát thấy trong rừng, nơi có rất ít ánh sáng mặt trời xuyên qua những tán cây [52],[102]. Véc tơ không có rừng (Non-forest vector) cho thấy sự thay đổi sinh thái học trong rừng. Ví dụ, An. culicifacies chủ yếu sống trong nhà [103] nhưng trong rừng rậm ở miền trung Ấn Độ, loài này được báo cáo chủ yếu sống ngoài nhà trong tự nhiên [104]. Tương tự như vậy, loài An. gambiae đã quan sát cho thấy sống chủ yếu ngoài nhà và thích nghi đốt người trong khu vực rừng so với khu vực không có rừng ở miền nam Sierra [55]. Chỉ số đốt người cao nhất và nhiễm thoa trùng đã được quan sát ở khu vực rừng xa van cho tất cả 4 véc tơ sốt rét chính gồm An. gambiae, An. funestus, An. arabiensis và An. moucheti ở Nigeria [105] và An. gambiae ở miền Nam Ethiopia [106] và Madagascar [107] so với các khu vực rừng nhiệt đới ít rừng ở phía tây nam Nigeria, nơi loài An. arabiensis đốt động vật chủ yếu, trong khi An. gambiae, An. melas và An. moucheti vẫn chủ yếu đốt người [108]. Các tác động của rừng/ phá rừng đối với các quần thể véc tơ, sinh thái học của véc tơ và tỷ lệ mắc bệnh sốt rét được tóm tắt trong Bảng 1. Bảng 1. Tác động của rừng/ phá rừng đối với các quần thể, sinh thái học véc tơ và tỷ lệ mắc sốt rét Véc tơ sốt rét | Gia tăng đốt người | Gia tăng sống ngoài nhà/thích đốt ngoài nhà | Gia tăng quần thể véc tơ/ sốt rét | An. gambiae | Vùng rừng của Nam Sierra [55] | Vùng rừng của nam Sierra [55] | Vùng rừng bị phá ở châu Phi [72],[109] | An. arabiensis | Vùng rừng Savan của Nigeria [105] | Vùng rừng của Nigeria [110] | Vùng rừng bị phá của châu Phi [72] | An. funestus | Vùng rừng Savan của Nigeria [105] | Không gia tăng tiêu máu ngoài nhà được báo cáo từ vùng rừng mưa của Nigeria [110] | Vùng rừng bị phá của Africa [72] | An. dirus | Vùng rừng của Thái Lan [52] và Việt Nam[17] | Vùng rừng của Việt Nam [17,28] | Vùng rừng của châu Á [72] | An. fluviatilis | Vùng rừng ở Orissa, Ấn Độ [111] | Vùng rừng của Trung Ấn Độ [104] | Vùng rừng của Orissa, Ấn Độ [111] | An. minimus | Vùng rừng của tỉnh Kratie, Campuchia [112] Vùng rừng bị phá của miền Trung Việt Nam [18] | Vùng rừng bị phá của miền Trung, Việt Nam[18] | Vùng rừng bị ohas của châu Á [72,79,80] | An. culicifacies | Vùng rừng của Orissa, Ấn Độ [111] | Vùng rừng của Trung Ấn Độ [104] | Vùng rừng bị phá của châu Á [72] | An. maculatus | Vùng rừng của tỉnh Kratie, Campuchia[112] | Vùng rừng của tỉnh Kratie, Campuchia [112] | Vùng rừng bị phá châu Á [72] | An. darlingi | Vùng rừng bị phá của Peru, Amazon [82] | Vùng rừng của Brazil [113] | Vùng rừng bị phá của Nam Mỹ [72],[82] và vùng rừng gần của Nam Mỹ [114] | An. aquasalis | Vùng rừng của Guayan, Venezuela [67] | Vùng rừng rụng lá của Venezuela [67] | Vùng rừng bị phá của Nam Mỹ [72]. |
Yếu tố ký sinh trùng liên quan đến véc tơ và vật chủ Phân bố các loài Plasmodium ở các vùng rừng Plasmodium ở người ít chịu ảnh hưởng bởi các yếu tố rừng, vì vòng đời thứ hai của nó hoàn thành ở động vật máu nóng. Tuy nhiên, chu kỳ sống chính xảy ra ở động vật máu lạnh là các véc tơ truyền bệnh do vậy chúng bị ảnh hưởng rất lớn bởi môi trường. Thời kỳ ủ bệnh bên trong được kích hoạt bởi các hiện tượng không rõ ràng [115] nhưng thời gian ủ bệnh bên ngoài tỷ lệ nghịch với nhiệt độ và cũng phụ thuộc vào các loài Plasmodium và véc tơ [116]. P. vivax và P. falciparum có thời gian ủ bệnh bên ngoài ngắn hơn và cũng là những loài ký sinh trùng sốt rét ở người phổ biến nhất [117]. P. vivax có thể tồn tại ở những nơi như miền Trung Andes, nơi An. pseudopunctipennis là véc tơ phụ và các điều kiện môi trường biến động bất thường thường xuyên xảy ra và được bù đắp bằng thời gian ủ bệnh P. vivax bên ngoài ngắn, thời gian ủ bệnh trong gan người lâu [118] và độ bao phủ rừng làm tăng tuổi thọ của các véc tơ sốt rét [109]. Các loài Plasmodium spp. đã tiến hóa để phù hợp với các véc tơ địa phương, như quan sát thấy loài P. falciparum ở các vùng nông thôn Cameroon bằng cách rút ngắn thời gian phát triển thoa trùng [119] vì sự sống còn của các véc tơ ảnh hưởng đến phân bố Plasmodium spp. [120]. Gia tăng khả năng của véc tơ An. gambiae đã được báo cáo ở các khu vực phá rừng của Kenya như phá rừng dẫn đến giảm thời gian hình thành bào tử của P. falciparum [109]. Các trường hợp nhiễm P. malariae, P. knowlesi và P. ovale rất hiếm và chủ yếu giới hạn trong khu vực rừng hẻo lánh và thường được báo cáo không đầy đủ vì thường xác định nhầm loài ký sinh trùng [121], [122],[123] cũng như các công cụ phân tử xác định loài không phải lúc nào cũng sẵn sàng tại các cơ sở nghiên cứu tại thực địa lẫn la bô chuyên sâu. Nguy cơ sốt rét động vật linh trưởng lây sang con người ở các vùng rừng Sự hiện diện các loài ký sinh trùng sốt rét Plasmodium spp. linh trưởng ở ổ bệnh trong rừng đặt ra nguy cơ lớn về sự lây lan ký sinh trùng sốt rét ở linh trưởng sang con người khi con người phá rừng, cũng như sự di chuyển ổ ạt của con người vào rừng [124]. Anopheles trong giống phụ Kerteszia (An Kerteszia cruzii., An. Kerteszia Bellator) là các véc tơ của con người và ký sinh trùng khỉ Plasmodia ở các vùng như rừng Đại Tây Dương ở Nam Mỹ [33],[34]. Điều đó có khả năng bệnh lây truyền từ động vật sang người có thể đã có mặt ở những khu vực này, do ký sinh trùng được tìm thấy ở các loài khỉ (P. simium và P. brasilianum) có cấu trúc di truyền tương tự hoặc liên quan đến ký sinh trùng Plasmodia ở người (P. vivax và P. malariae) [35]. Những trường hợp như vậy đã xảy ra, mặc dù không thường xuyên trong rừng hoặc bìa rừng, mà đặc điểm của ký sinh trùng động vật linh trưởng có thể bị nhầm lẫn với hình thái tương tự như ký sinh trùng plasmodia ở người [33]. Các loài Plasmodium ở khỉ có thể đã lây sang con người như đã xảy ra trong trường hợp P. cynomolgi ở Ấn Độ [125], P. simium ở Brazil [126],[127], P. knowlesi ở Malaysia [20.124]. Sự có mặt những cá nhân bị ký sinh trùng không có triệu chứng cùng với khỉ bị nhiễm bệnh có thể duy trì lan truyền bệnh sốt rét tự nhiên ở nhưng nơi mà giám sát và phòng chống bệnh sốt rét khó khăn [33,36]. Nguồn chứa ký sinh trùng và kháng thuốc Sự lan truyền bệnh sốt rét ổn định ở các ùng lưu hành cũng được báo cáo là có liên quan đến những người nhiễm P. falciparum và P. vivax không có biểu hiện lâm sàng và các ổ chứa P. vivax thể ẩn hay thể ngủ ở trong gan (hypnozoite form) [128],[129]. Sốt rét không có triệu chứng thường gắn liền với các khu vực có rừng trên thế giới [130],[133]. Do các ổ chứa ký sinh trùng không có triệu chứng [131], sốt rét lưu hành ổn định được duy trì liên tục trong các khu vực rừng [134] và ở các khu vực không có rừng với môi trường không ổn định thì các ổ chứa ký sinh trùng đóng một vai trò rất quan trọng trong việc chuyển tiếp các mùa lan truyền bệnh [134]. Chẳng hạn, lan truyền luôn tái diễn mạnh mẽ thông qua một ổ chứa ký sinh trùng sốt rét không có triệu chứng lâm sàng đã được báo cáo ở các khu vực ven sông có rừng của Tanzania [131] và Gabon [130]. Bệnh nhân nhiễm ký sinh trùng sốt rét nhưng không có triệu chứng lâm sàng [129, 135] được xem như ổ chứa [132],[136],[137] và sẵn sàng là nguồn lây nhiễm cho các véc tơ [136], do vậy đây là một trong những trở ngại tiềm ẩn để kiểm soát bệnh sốt rét trong rừng. Bệnh nhân nhiễm Plasmodium spp. kháng thuốc mang giao bào lây truyền trong một thời gian rất dài và hoạt động như các ổ chứa chủng kháng thuốc. Loài P. falciparum và P. vivax được báo cáo là thường xuyên liên quan đến kháng thuốc tại các khu rừng nơi mà cường độ lan truyền sốt rét cao hơn và kiểm soát bệnh sốt rét thường bị bỏ quên. Vì những lý do này, khu vực biên giới Thái Lan - Campuchia có rừng được cho là "cái nôi" về nguồn gốc của kháng chloroquin và phát triển đa kháng thuốc [24],[30],[31]. Gần đây, một phần kháng artemisinin do P. falciparum đã nổi lên từ những khu vực này [24],[32]. Việc phổ biến chủng ký sinh trùng kháng thuốc nhanh hơn có thể do sự có mặt một số các véc tơ rừng rất hiệu quả ở khu vực Đông Nam Á như An. dirus và An. minimus, các véc tơ này cho thấy lần lượt 66% và 44% nhạy cảm với chủng P. falciparum kháng thuốc [28],[52] và số lượng nang trứng của loài ký sinht rùng kháng thuốc được báo cáo cao hơn ở An. dirus [52]. Trung tâm kháng chloroquin cũng đã được tìm thấy liên quan đến muỗi An. dirus ở Nam Á [52],[138]. Các yếu tố sinh thái và kinh tế xã hội của con người Các thực hành và tập quán liên quan đến sốt rét của quần thể dân tộc bản địa Khu vực rừng sâu chủ yếu là nơi sinh sống các cộng đồng dân tộc thiểu số bản địa và các bộ lạc [47],[48], họ rất ít liên lạc với thế giới bên ngoài và chủ yếu phụ thuộc vào rừng để nuôi sống bản thân và các thành viên gia đình [139]. Do vậy các cộng đồng hầu như mù chữ, dễ bị mê tín và ít giao tiếp với nhân viên phòng chống bệnh sốt rét. Những bộ lạc sống trong rừng đã được bảo tồn truyền thống và thói quen có tác động đáng kể đến lan truyền bệnh sốt rét [17], do vậy những người sống trong rừng này rất dễ bị sốt rét [21]. Đốt nương làm rẫy là một yếu tố chính của tập quán canh tác các cộng đồng dân tộc thiểu số bản địa ở khu vực rừng nhiều nơi trên thế giới [17],[140] điều này dẫn đến nạn phá rừng, thay đổi môi trường sinh thái rừng, do đó làm thay đổi mô hình lan truyền sốt rét [17]. Loại hình canh tác này thường có 1 - 2 thành viên trong gia đình ngủ qua đêm trong nhà rẫy, những người này gia tăng nguy cơ bị véc tơ sốt rét đốt [17],[140]. Ví dụ, các điều kiện thuận lợi được tạo ra ở khu vực rừng núi miền Trung Việt Nam, nơi tập quán của cộng đồng dân tộc RagLay đốt rừng làm nương rẫy [17[,[140] và cũng do khai thác gỗ thương mại và chăn nuôi gia súc [141]. Một số cộng đồng dân tộc ở Ấn Độ theo nghi lễ bôi đầy phân bò tươi và bùn lên ngôi nhà của họ làm phủ kín hóa chất diệt côn trùng đã phun tồn lưu trên tường vách và làm cho nó không hiệu quả để kiểm soát véc tơ [142]. Theo truyền thống, người phụ nữ phủ kín cơ thể và có lẽ đây là lý do làm cho họ có nguy cơ sốt rét thấp hơn so với nam giới [17],[61],[143]. Ở nhiều bộ lạc cả nam giới và phụ nữ đều uống rượu và các sản phẩm có cồn thường xuyên và đây là nguyên nhân làm giảm sự tự bảo vệ cá nhân chống lại muỗi đốt. Điều thú vị đã được tìm thấy rằng, tiêu thụ bia làm tăng sức hấp dẫn con người đối với An. gambiae trong các thí nghiệm được tiến hành ở Burkina Faso [144]. Quần thể dân cư đi vào rừng Người dân di chuyển vào trong và ra khỏi rừng vì nhiều lý do khác nhau, điều này giúp lan truyền sốt rét được duy trì [145],[146]. Khoảng cách di chuyển ngắn hàng ngày để chăn thả gia súc, săn bắn, đánh cá, trồng trọt, thu lâm sản như lá, gỗ, cây ăn quả, hoa và mật ong,… [147]. Như vây, sự đi lại làm gia tăng tiếp xúc giữa người với véc tơ sốt rét nhất là ban đêm ngủ lại ở trong rừng [145],[148],[49] và ngay cả vào ban ngày có ánh sáng nơi véc tơ như An. dirus chiếm ưu thế [52.140]. Cộng đồng dân tộc sống đi lại với một khoảng cách ngắn đến trung bình ở trong rừng được thấy trong thời gian gieo cấy và các mùa thu hoạch [38]. Nói chung, sự di chuyển này mang sốt rét từ rừng ra những khu vực đồng bằng [145]. Ví dụ, sự gia tăng đi lại của người dân ở cả trong các vùng cao nguyên New Guinea và cũng như giữa các vùng đất thấp lưu hành sốt rét cao và vùng cao nguyên đã làm gia tăng lan truyền sốt rét [150]. Những cư dân sống ngoài rừng cũng đi vào các khu vực rừng để chăn thả gia súc và thu gỗ [151]. Người tị nạn đã được giải quyết và nhiều chương trình tái định cư đã được đưa ra ở các khu vực rừng, ví dụ, ở Ấn Độ, người tị nạn Bangladesh được chuyển sang huyện Chittagong với nhiều đồi, rừng và khu vực rừng biên giới Orissa - Chhattisgarh gần Bastar thuộc dự án ‘Dandakaranya’. Những người tị nạn bị bệnh sốt rét ở các bộ tộc bản địa, điều này dẫn đến dịch bệnh ở Bastar [15],[21],[53]. Theo Lindsay và cộng sự, "Nhiều người trong số những người định cư châu Âu đầu tiên ở châu Phi đã tìm cách lánh nạn từ vùng nóng và các bệnh của vùng đồng bằng lớn di chuyển đến vùng cao nguyên có khí hậu mát mẻ và trong lành cũng mang bệnh sốt rét" [154]. Truyền thông và cơ sở hạ tầng yếu kém Cộng đồng sống trong rừng thường xây dựng những ngôi nhà bằng bùn và nhiễm bệnh gia tăng trong số những người sống trong các ngôi nhà bằng bùn hoặc những ngôi nhà được xây dựng tạm bợ gần nơi sinh sản của véc tơ sốt rét ở Ai Cập [155], Ethiopia [156] và Kenya [42]. An. minimus A ở miền Trung Việt Nam cho thấy tỷ lệ hoạt động trong nhà và ưa đốt người cao, rất có thể bị ảnh hưởng bởi những ngôi nhà có cấu trúc tường vách không kín với nhiều khe hở thuận lợi cho việc phát hiện con người và xâm nhập vào nhà dân [157]. Cơ sở hạ tầng y tế và giám sát đang bị bỏ quên ở những khu vực rừng núi xa xôi [158] và điều này trở nên khó khăn hơn vào mùa mưa, nơi cơ sở hạ tầng đường bộ thiếu thốn hoặc không có [159]. Nhưng nghiêm trọng hơn, mùa mưa cũng là khoảng thời gian lan truyền bệnh cao điểm ở hầu hết các vùng dịch tễ sốt rét [160]. Khi thực hiện giám sát đầy đủ theo yêu cầu thì những bệnh nhân phát hiện cảm thấy rất khó khăn để tiếp cận cơ sở y tế do khí hậu và các vấn đề giao tiếp [21,45]. Do đó, người dân bản địa nói chung thường dựa trên các hoạt động y tế của những thầy địa phương và / hoặc thầy lang [161-163]. Báo cáo cho biết rằng thói quen tìm kiếm điều trị tỷ lệ nghịch với khoảng cách từ một cơ sở y tế và các vấn đề truyền thông [21,165,166]. Sự hợp tác và quan niệm của người dân Sự nhận thức của cộng đồng các bộ tộc về bệnh sốt rét đã được báo cáo là khác nhau ở nhiều nơi trên thế giới. Một số bộ lạc tin rằng bệnh sốt rét là do thần linh, do vị thần nổi giận, do tà ma hoặc có nơi xem đó là một cơn sốt tự giới hạn ở các nước như Ấn Độ [167] và Tây Nam Ethiopia [165]. Chi phí điều trị bệnh thấp với các loại thuốc truyền thống, khả năng tiếp cận tốt và giao tiếp tốt với các thầy lang được ưa thích nhất trong các khu vực rừng hẻo lánh, xa các trung tâm y tế của chính phủ như đã báo cáo ở vùng nông thôn Ethiopia [168]. Trong các khu vực rừng, việc tìm kiếm chăm sóc sức khỏe có liên quan trực tiếp đến văn hóa, niềm tin và khả năng chi trả các dịch vụ chăm sóc sức khỏe [21, 165]. Điều đó cho thấy rằng "các dịch vụ y tế có thể được sử dụng đúng mức và một số hướng dẫn chăm sóc sức khỏe có thể không có hiệu quả hoặc bị bỏ qua trong các xã hội truyền thống và xã hội chuyển tiếp nơi mà những ý tưởng của người dân và các hành vi xung đột với những kiến thức được truyền cho họ" [161]. Nói chung nghèo đói là yếu tố quan trọng nhất trong việc tiếp cận cơ sở y tế bên cạnh đó mù chữ, mê tín dị đoan, và đức tin văn hóa giữa các dân tộc bản địa ở hầu hết các vùng rừng Bangladesh [143]. Cư dân sống trong các khu rừng thường là nghèo, không có màn ngủ và nếu có màn thì giá cả cao nên không thể tiếp cận mặc dù thực tế người dân biết những lợi ích của màn ngủ và muốn sử dụng chúng [169, 170]. Do không có màn ngủ nên người nghèo không bảo vệ cơ thể tránh bị muỗi đốt và do thiếu cơ sở y tế hiện đại với giá cả phải chăng vì vậy họ thực hiện các biện pháp truyền thống [171]. Giám sát không đạt yêu cầu ở các vùng rừng núi xa xôi [172] và thuốc sốt rét điều trị ở liều thấp được thực hiện cho tất cả các loại ký sinh trùng sốt rét [173], chính điều này thúc đẩy sự phát triển kháng thuốc [174,175]. Như vậy, nguồn gốc và sự phát triển kháng thuốc đã được báo cáo đầu tiên ở các vùng rừng xa xôi Campuchia và khu vực biên giới Thái Lan-Myanmar [176]. Khắc phục thách thức chính trong việc phòng chống bệnh sốt rét ở các hệ sinh thái rừng Những nỗ lực kiểm soát sốt rét rừng không đạt yêu cầu chủ yếu là do truyền thông kém và đường xá đi lại khó khăn [21]. Phát triển cơ sở hạ tầng truyền thông phải được ưu tiên hàng đầu vì điều này sẽ mở ra những cách thức và cơ hội mới cũng như thúc đẩy phát triển kinh tế-xã hội cho người dân. Thành lập và phát triển các cơ sở y tế để người dân có thể tiếp cận dịch vụ dễ dàng bằng cách giảm khoảng cách giữa các địa phương và nhân viên y tế có thể cải thiện được tình hình. Nâng cao nhận thức xã hội để kiểm soát bệnh sốt rét và sự tham gia của các nhân viên y tế truyền thống và phi chính phủ có thể giúp đỡ trong việc khắc phục những khoảng trống trong các khu vực rừng lạc hậu [21],[162]. Lan truyền bệnh sốt rét không có ranh giới chính trị và ở các khu vưc biên giới rừng con người thường di chuyển qua lại biên giới và mang bệnh sốt rét như đã thấy ở biên giới Thái Lan-Myanmar. Sự di dân ở các vùng rừng có nhiều nguyên nhân khác nhau nhưng chủ yếu là do có sẵn của công trình [49],[97]. Doanh trại quân đội, các trạm radar, cảnh sát và lực lượng vũ trang cấm trại trại khác và các dự án phát triển lớn như đường bộ và phát triển cơ sở hạ tầng khác, khai thác khoáng sản, hoạt động nông nghiệp như chè, cao su, trồng cà phê và xây dựng các đập nước sử dụng số lượng lớn lao động nhập cư. Những người này dễ mắc sốt rét từ người dân địa phương [152]. Những người di cư và người tị nạn không được bảo vệ cần phải được kiểm tra và điều trị kịp thời đối với bệnh sốt rét. Quân đội và các trại khác trong khu vực rừng được yêu cầu phải có những nỗ lực lớn để chiến đấu chống lại bệnh sốt rét cùng với người dân địa phương. Các bộ lạc trong vùng rừng thường không tiếp xúc với thế giới bên ngoài, nói chung họ thường bảo thủ và miễn cưỡng trong việc tìm kiếm điều trị. Những hạn chế xã hội, sự thiếu hiểu biết, mê tín dị đoan và sự chịu đựng của bản thân khi mang bệnh sốt rét có triệu chứng hoặc không có triệu chứng, những người này không tìm cách điều trị và hành động như là các ổ chứa ký sinh trùng sốt rét [21],[143]. Sàng lọc đại trà có thể hữu ích để đánh giá bệnh sốt rét bằng phương pháp huyết thanh dương tính trong số các cộng đồng nơi mà không có triệu chứng sốt rét chiếm ưu thế và mọi người ít tìm kiếm điều trị [23]. Kháng nguyên dựa vào bộ dụng cụ xét nghiệm chẩn đoán nhanh đặc hiệu loài nên được sử dụng trong các khu vực rừng để phát hiện và điều trị bệnh tại chỗ. Các Bộ lạc du mục nguyên thủy cần phải được tiếp cận bởi các nhà cung cấp chăm sóc sức khỏe và cần phải được khuyến khích sử dụng y tế và các dịch vụ khác. Rừng được báo cáo là trung tâm của sự lây lan kháng thuốc sốt rét và đánh giá kết quả trong kiểm soát bệnh sốt rét thấp. Vì vậy, quản lý kiểm soát chặt chẽ thuốc sốt rét với đánh giá định kỳ về tình trạng kháng thuốc được đề xuất trong các khu vực rừng. Các Marker phân tử liên quan đến kháng thuốc sốt rét cần phải được đánh giá ở các khu vực rừng lan truyền cao và có thể giúp tiết kiệm các thuốc sốt rét có giá trị cho thế hệ sau [176]. Phá rừng và biến đổi khí hậu có thể gọp vào với các yếu tố khác của sốt rét rừng gây ra một sự bùng nổ các loài véc tơ và sự gia tăng quy mô véc tơ [20],[82]. Sự nối tiếp sinh thái của các véc tơ sốt rét do sự thay đổi khí hậu và sinh thái cần được tìm hiểu và kiểm tra thường xuyên sự phong phú của các loài véc tơ và cập nhật về mô hình phân bố, sinh thái học và thay đổi hành vi ở khu vực có rừng là cơ sở cho các biện pháp kiểm soát véc tơ có hiệu quả. Thay lời kết Sự tồn tại mối liên quan chặc chẽ giữa các yếu tố khác nhau ảnh hưởng đến động lực truyền bệnh sốt rét ở các hệ sinh thái rừng. Một thay đổi nhỏ trong bất kỳ các yếu tố nào đều ảnh hưởng đến những yếu tố khác, mà đỉnh cao là mô hình lan truyền khác nhau. Thay đổi độ bao phủ thảm thực vật và phá rừng làm thay đổi sự phân bố và tập tính của các véc tơ sốt rét. Đặc điểm sinh thái và kinh tế xã hội con người cũng ảnh hưởng đến tập tính ưa đốt mồi ngoài nhà, hoạt động ngoài nhà, thời gian đốt và hành vi trú đậu của véc tơ. Các đặc điểm di truyền của các cộng đồng sống tại các khu rừng và thói quen tìm kiếm chăm sóc sức khỏe của họ là rất quan trọng đối với sự phổ biến ký sinh trùng và phát triển kháng thuốc. Ngoài ra, sự đi lại và các hoạt động ở trong rừng là yếu tố quan trọng liên quan đến mô hình lan truyền bệnh sốt rét. Do vậy, lan truyền sốt rét ở các khu vực rừng là quá trình phức tạp liên quan đến sự tác động qua lại giữa các yếu tố địa hình, côn trùng, ký sinh trùng và con người. Các nghiên cứu đã thực hiện ở các khu vực rừng tại nhiều vùng khác nhau trên thế giới nhìn chung tập trung vào một hoặc một vài trong nhiều yếu tố mà điều này có thể không đầy đủ để hiểu biết sự phức tạp của tình hình sốt rét phát sinh do sự tương tác một vài yếu tố. Phần đánh giá này có thể giúp hiểu biết sự phức tạp liên kết giữa các yếu tố khác nhau tác động đồng thời ảnh hưởng đến đến lan truyền sốt rét. Mô hình dự báo lan truyền sốt rét có thể được thực hiện đối với các khu vực rừng của các vùng sinh thái khác nhau bằng cách đưa vào phát hiện tất cả các yếu tố điều chỉnh có liên quan. Dựa vào sự hiểu biết rõ ràng về mối quan hệ phức tạp của các thông số khác nhau, tình hình chiến lược kiểm soát sốt rét/véc tơ cụ thể có thể được phát triển và thực hiện để giải quyết vấn đề sốt rét ở rừng hiện nay. Mặc dù thực hiện chiến lược này là trách nhiệm chính của chính phủ và các cơ quan y tế địa phương, các tổ chức phi chính phủ, các bác sĩ y tế địa phương, nhưng những người chữa bệnh truyền thống sẽ là quan trọng vì đây là sự chấp nhận đối với các cộng đồng sinh sống trong vùng rừng xa xôi, hẻo lánh. Đồng thời thiết lậpmối quan hệ tốt thông qua sự tương tác giữa người thực hiện và cộng đồng là điều cần thiết cho sự thành công và hỗ trợ các chương trình phòng chống sốt rétở các hệ sinh thái rừng. Tài liệu tham khảo 1.WHO: World Malaria Report: 2013. Switzerland: World Health Organization; 2013:1-199. 2.Gething PW, Patil AP, Smith DL, Guerra CA, Elyazar IR, Johnston GL, Tatem AJ, Hay SI: A new world malaria map: Plasmodium falciparum endemicity in 2010. Malar J 2011, 10:378. 3.Gething PW, Elyazar IR, Moyes CL, Smith DL, Battle KE, Guerra CA, Patil AP, Tatem AJ, Howes RE, Myers MF, George DB, Horby P, Wertheim HF, Price RN, Mueller I, Baird JK, Hay SI: A long neglected world malaria map: Plasmodium vivax endemicity in 2010. PLoS Negl Trop Dis 2012, 6:e1814. 4.Collins WE, Jeffery GM: Plasmodium malariae: parasite and disease. Clin Microbiol Rev 2007, 20:579-592. 5.Obare P, Ogutu B, Adams M, Odera JS, Lilley K, Dosoo D, Adhiambo C, Owusu-Agyei S, Binka F, Wanja E, Johnson J: Misclassification of Plasmodium infections by conventional microscopy and the impact of remedial training on the proficiency of laboratory technicians in species identification. Malar J 2013, 12:113. 6.Sutherland CJ, Tanomsing N, Nolder D, Oguike M, Jennison C, Pukrittayakamee S, Dolecek C, Hien TT, Do Rosario VE, Arez AP, Pinto J, Michon P, Escalante AA, Nosten F, Burke M, Lee R, Blaze M, Otto TD, Barnwell JW, Pain A, Williams J, White NJ, Day NP, Snounou G, Lockhart PJ, Chiodini PL, Imwong M, Polley SD: Two nonrecombining sympatric forms of the human malaria parasite Plasmodium ovale occur globally. J Infect Dis 2010, 201:1544-1550. 7.White NJ: Plasmodium knowlesi: the fifth human malaria parasite. Clin Infect Dis 2008, 46:172-173. 8.Putaporntip C, Hongsrimuang T, Seethamchai S, Kobasa T, Limkittikul K, Cui L, Jongwutiwes S: Differential prevalence of Plasmodium infections and cryptic Plasmodium knowlesi malaria in humans in Thailand. J Infect Dis 2009, 199:1143-1150. 9.Singh B, Kim Sung L, Matusop A, Radhakrishnan A, Shamsul SS, Cox-Singh J, Thomas A, Conway DJ: A large focus of naturally acquired Plasmodium knowlesi infections in human beings. Lancet 2004, 363:1017-1024. 10.Van den Eede P, Van HN, Van Overmeir C, Vythilingam I, Duc TN, Hung Le X, Manh HN, Anne J, D’Alessandro U, Erhart A: Human Plasmodium knowlesi infections in young children in central Vietnam. Malar J 2009, 8:249. 11.Vythilingam I, Noorazian YM, Huat TC, Jiram AI, Yusri YM, Azahari AH, Norparina I, Noorrain A, Lokmanhakim S: Plasmodium knowlesi in humans, macaques and mosquitoes in peninsular Malaysia. Parasit Vectors 2008, 1:26. 12.Guerra CA, Snow RW, Hay SI: A global assessment of closed forests, deforestation and malaria risk. Ann Trop Med Parasitol 2006, 100:189-204. 13.Snelder DJ, Lasco RD, NetLibrary I: Smallholder Tree Growing for Rural Development and Environmental Services Lessons from Asia. Dordrecht: Springer; 2008:1-493. 14.Breckle S-W, Walter H: Walter’s Vegetation of the Earth: The Ecological Systems of the Geo Biosphere. Berlin [u.a.]: Springer; 2002:1-236. 15.Achard F, Eva HD, Stibig HJ, Mayaux P, Gallego J, Richards T, Malingreau JP: Determination of deforestation rates of the world’s humid tropical forests. Science 2002, 297:999-1002. 16.Mayaux P, Holmgren P, Achard F, Eva H, Stibig HJ, Branthomme A: Tropical forest cover change in the 1990s and options for future monitoring. Philos Trans R Soc Lond B Biol Sci 2005, 360:373-384. 17.Erhart A, Ngo DT, Phan VK, Ta TT, Van Overmeir C, Speybroeck N, Obsomer V, Le XH, Le KT, Coosemans M, D’Alessandro U: Epidemiology of forest malaria in central Vietnam: a large scale cross-sectional survey. Malar J 2005, 4:58. 18.Trung HD, Bortel WV, Sochantha T, Keokenchanh K, Briet OJ, Coosemans M: Behavioural heterogeneity of Anopheles species in ecologically different localities in Southeast Asia: a challenge for vector control. Trop Med Int Health 2005, 10:251-262. 19.Walker M, Winskill P, Basanez MG, Mwangangi JM, Mbogo C, Beier JC, Midega JT: Temporal and micro-spatial heterogeneity in the distribution of Anopheles vectors of malaria along the Kenyan coast. Parasit Vectors 2013, 6:311. 20.Alias H, Surin J, Mahmud R, Shafie A, Mohd Zin J, Mohamad Nor M, Ibrahim AS, Rundi C: Spatial distribution of malaria in Peninsular Malaysia from 2000 to 2009. Parasit Vectors 2014, 7:186. 21.Ribera JM, Hausmann-Muela S: The straw that breaks the camel’s back. Redirecting health-seeking behavior studies on malaria and vulnerability. Med Anthropol Q 2011, 25:103-121. 22.Tatem AJ, Smith DL, Gething PW, Kabaria CW, Snow RW, Hay SI: Ranking of elimination feasibility between malaria-endemic countries. Lancet 2010, 376:1579-1591. 23.Hamainza B, Moonga H, Sikaala CH, Kamuliwo M, Bennett A, Eisele TP, Miller J, Seyoum A, Killeen GF: Monitoring, characterization and control of chronic, symptomatic malaria infections in rural Zambia through monthly household visits by paid community health workers. Malar J 2014, 13:128. 24.Salud OM: Global Malaria Control and Elimination: Report of a Meeting on Containment of Artemisinin Tolerance, 19 January 2008, Geneva, Switzerland. Geneva: WHO; 2008:1-29. 25.Hay SI, Guerra CA, Tatem AJ, Noor AM, Snow RW: The global distribution and population at risk of malaria: past, present, and future. Lancet Infect Dis 2004, 4:327-336. 26.Carter R, Mendis KN: Evolutionary and historical aspects of the burden of malaria. Clin Microbiol Rev 2002, 15:564-594. 27.Fuller DO, Parenti MS, Hassan AN, Beier JC: Linking land cover and species distribution models to project potential ranges of malaria vectors: an example using Anopheles arabiensis in Sudan and Upper Egypt. Malar J 2012, 11:264. 28.Trung HD, Van Bortel W, Sochantha T, Keokenchanh K, Quang NT, Cong LD, Coosemans M: Malaria transmission and major malaria vectors in different geographical areas of Southeast Asia. Trop Med Int Health 2004, 9:230-237. 29.Charlwood JD, Vij R, Billingsley PF: Dry season refugia of malaria-transmitting mosquitoes in a dry savannah zone of east Africa. Am J Trop Med Hyg 2000, 62:726-732. 30.Guthmann JP, Pittet A, Lesage A, Imwong M, Lindegardh N, Min Lwin M, Zaw T, Annerberg A, De Radigues X, Nosten F: Plasmodium vivax resistance to chloroquine in Dawei, southern Myanmar. Trop Med Int Health 2008, 13:91-98. 31.Nosten F, ter Kuile F, Chongsuphajaisiddhi T, Luxemburger C, Webster HK, Edstein M, Phaipun L, Thew KL, White NJ: Mefloquine-resistant falciparum malaria on the Thai-Burmese border. Lancet 1991, 337:1140-1143. 32.Dondorp AM, Yeung S, White L, Nguon C, Day NP, Socheat D, von Seidlein L: Artemisinin resistance: current status and scenarios for containment. Nat Rev Microbiol 2010, 8:272-280. 33.Duarte AM, Pereira DM, de Paula MB, Fernandes A, Urbinatti PR, Ribeiro AF, Mello MH, Matos MO Jr, Mucci LF, Fernandes LN, Natal D, Malafronte RS: Natural infection in anopheline species and its implications for autochthonous malaria in the Atlantic Forest in Brazil. Parasit Vectors 2013, 6:58. 34.Deane LM, Ferreira Neto JA, Lima MM: The vertical dispersion of Anopheles (Kerteszia) cruzi in a forest in southern Brazil suggests that human cases of malaria of simian origin might be expected. Mem Inst Oswaldo Cruz 1984, 79:461-463. 35.Waters AP, Higgins DG, McCutchan T: Evolutionary relatedness of some primate models of Plasmodium. Mol Biol Evol 1993, 10:914-923. 36.De Castro Duarte AMR, Malafronte RS, Cerutti C Jr, Curado I, De Paiva BR, Maeda AY, Yamasaki T, Summa MEL, Neves DVDA, De Oliveira SG: Natural Plasmodium infections in Brazilian wild monkeys: reservoirs for human infections? Acta Trop 2008, 107:179-185. 37.Rosenberg R: Forest malaria in Bangladesh. III. Breeding habits of Anopheles dirus. Am J Trop Med Hyg 1982, 31:192-201. 38.Rubio-Palis Y, Zimmerman RH: Ecoregional classification of malaria vectors in the neotropics. J Med Entomol 1997, 34:499-510. 39.Minakawa N, Sonye G, Mogi M, Githeko A, Yan G: The effects of climatic factors on the distribution and abundance of malaria vectors in Kenya. J Med Entomol 2002, 39:833-841. 40.Koenraadt CJ, Githeko AK, Takken W: The effects of rainfall and evapotranspiration on the temporal dynamics of Anopheles gambiae s.s. and Anopheles arabiensis in a Kenyan village. Acta Trop 2004, 90:141-153. 41.Aragao MB: Some microclimatic measurements, in a forest of the “bromeliad-malaria” region, in Santa Catarina, Brazil. III. Conditions of humidity measured with hygrographs. Rev Bras Med 1960, 12:395-414. 42.Zhou G, Munga S, Minakawa N, Githeko AK, Yan G: Spatial relationship between adult malaria vector abundance and environmental factors in western Kenya highlands. Am J Trop Med Hyg 2007, 77:29-35. 43.Manda H, Gouagna LC, Foster WA, Jackson RR, Beier JC, Githure JI, Hassanali A: Effect of discriminative plant-sugar feeding on the survival and fecundity of Anopheles gambiae. Malar J 2007, 6:113. 44.Okech BA, Gouagna LC, Yan G, Githure JI, Beier JC: Larval habitats of Anopheles gambiae s.s. (Diptera: Culicidae) influences vector competence to Plasmodium falciparum parasites. Malar J 2007, 6:50. 45.Lim ES: Current status of malaria in Malaysia. Southeast Asian J Trop Med Public Health 1992, 23(Suppl 4):43-49. 46.Rattanarithikul R, Green CA, Panyim S, Noigamol C, Chanaimongkol S, Mahapibul P: Larval habitats of malaria vectors and other Anopheles mosquitoes around a transmission focus in northwestern Thailand. J Am Mosq Control Assoc 1995, 11:428-433. 47.Mandal H, Mukherjee S, Datta A, India AS: India, an Illustrated Atlas of Tribal World. Kolkata: Anthropological Survey of India Calcutta, Ministry of Tourism and Culture, Dept. of Culture, Govt. of India; 2002:1-106. 48.Kusel J, Adler E: Forest Communities, Community Forests. Lanham, Md.: Rowman & Littlefield Publishers; 2003:1-276. 49.Pichainarong N, Chaveepojnkamjorn W: Malaria infection and life-style factors among hilltribes along the Thai-Myanmar border area, northern Thailand. Southeast Asian J Trop Med Public Health 2004, 35:834-839. 50.Nath DC, Mwchahary DD: Association between climatic variables and malaria incidence: a study in Kokrajhar district of Assam, India. Glob J Health Sci 2013, 5:90-106. 51.Kristan M, Abeku TA, Beard J, Okia M, Rapuoda B, Sang J, Cox J: Variations in entomological indices in relation to weather patterns and malaria incidence in East African highlands: implications for epidemic prevention and control. Malar J 2008, 7:231. 52.Obsomer V, Defourny P, Coosemans M: The Anopheles dirus complex: spatial distribution and environmental drivers. Malar J 2007, 6:26. 53.Carnevale P, Le Goff G, Toto JC, Robert V: Anopheles nili as the main vector of human malaria in villages of southern Cameroon. Med Vet Entomol 1992, 6:135-138. 54.Wanji S, Tanke T, Atanga SN, Ajonina C, Nicholas T, Fontenille D: Anopheles species of the mount Cameroon region: biting habits, feeding behaviour and entomological inoculation rates. Trop Med Int Health 2003, 8:643-649. 55.Bockarie MJ, Service MW, Toure YT, Traore S, Barnish G, Greenwood BM: The ecology and behaviour of the forest form of Anopheles gambiae s.s. Parassitologia 1993, 35(Suppl):5-8. 56.Jawara M, Pinder M, Drakeley CJ, Nwakanma DC, Jallow E, Bogh C, Lindsay SW, Conway DJ: Dry season ecology of Anopheles gambiae complex mosquitoes in The Gambia. Malar J 2008, 7:156. 57.Mouchet J, Carnevale P: Impact of changes in the environment on vector-transmitted diseases. Sante 1997, 7:263-269. 58.Pavlovsky EN: Natural Nidality of Transmissible Diseases: With Special Reference to the Landscape Epidemiology of Zooanthroponoses. Urbana: University of Illinois Press; 1966:1-261. 59.O’Loughlin SM, Okabayashi T, Honda M, Kitazoe Y, Kishino H, Somboon P, Sochantha T, Nambanya S, Saikia PK, Dev V, Walton C: Complex population history of two Anopheles dirus mosquito species in Southeast Asia suggests the influence of Pleistocene climate change rather than human-mediated effects. J Evol Biol 2008, 21:1555-1569. 60.Hakre S, Masuoka P, Vanzie E, Roberts DR: Spatial correlations of mapped malaria rates with environmental factors in Belize. Central America Int J Health Geogr 2004, 3:6. 61.Haque U, Sunahara T, Hashizume M, Shields T, Yamamoto T, Haque R, Glass GE: Malaria prevalence, risk factors and spatial distribution in a hilly forest area of Bangladesh. PLoS one 2011, 6:e18908. 62.Afrane YA, Zhou G, Lawson BW, Githeko AK, Yan G: Effects of microclimatic changes caused by deforestation on the survivorship and reproductive fitness of Anopheles gambiae in western Kenya highlands. Am J Trop Med Hyg 2006, 74:772-778. 63.Yadav RS, Sharma VP, Chand SK: Mosquito breeding and resting in treeholes in a forest ecosystem in Orissa. Indian J Malariol 1997, 34:8-16. 64.Gunasekaran K, Sahu SS, Parida SK, Sadanandane C, Jambulingam P, Das PK: Anopheline fauna of Koraput district, Orissa state, with particular reference to transmission of malaria. Indian J Med Res 1989, 89:340-343. 65.Gary RE Jr, Foster WA: Anopheles gambiae feeding and survival on honeydew and extra-floral nectar of peridomestic plants. Med Vet Entomol 2004, 18:102-107. 66.Okech BA, Gouagna LC, Killeen GF, Knols BG, Kabiru EW, Beier JC, Yan G, Githure JI: Influence of sugar availability and indoor microclimate on survival of Anopheles gambiae (Diptera: Culicidae) under semifield conditions in western Kenya. J Med Entomol 2003, 40:657-663. 67.Berti J, Zimmerman R, Amarista J: Adult abundance, biting behavior and parity of Anopheles aquasalis, Curry 1932 in two malarious areas of Sucre State, Venezuela. Mem Inst Oswaldo Cruz 1993, 88:363-369. 68.Lindsay S: Environmental Management for Malaria Control in the East Asia and Pacific (EAP) Region. Washington: World Bank; 2004:1-46. 69.Kobayashi J, Somboon P, Keomanila H, Inthavongsa S, Nambanya S, Inthakone S, Sato Y, Miyagi I: Malaria prevalence and a brief entomological survey in a village surrounded by rice fields in Khammouan province, Lao PDR. Trop Med Int Health 2000, 5:17-21. 70.Sahu SS, Parida SK, Sadanandane C, Gunasekaran K, Jambulingam P, Das PK: Breeding habitats of malaria vectors: A. fluviatilis, A. annularis and A. culicifacies, in Koraput district, Orissa. Indian J Malariol 1990, 27:209-216. 71.Tuno N, Okeka W, Minakawa N, Takagi M, Yan G: Survivorship of Anopheles gambiae sensu stricto (Diptera: Culicidae) larvae in western Kenya highland forest. J Med Entomol 2005, 42:270-277. 72.Yasuoka J, Levins R: Impact of deforestation and agricultural development on anopheline ecology and malaria epidemiology. Am J Trop Med Hyg 2007, 76:450-460. 73.Sharma VP, Prasittisuk C, Kondrashin AV: Forest Malaria in Southeast Asia. In Proceedings of an Informal Consultative Meeting WHO/MRC; New Delhi. Edited by Sharma VP, Kondrashin AV. New Delhi: Malaria Research Centre; 1991:29-53. 74.Service MW: Agricultural development and arthropod-borne diseases: a review. Rev Saude Publica 1991, 25:165-178. 75.Service MW: Demography and Vector-Borne Diseases. Boca Raton: CRC Press; 1989:1-416. 76.Gilles HM, Warrell DA: Bruce-Chwatt’s Essential Malariology. London: A Hodder Arnold Publication; 1999:1-348. 77.Vittor AY, Pan W, Gilman RH, Tielsch J, Glass G, Shields T, Sanchez-Lozano W, Pinedo VV, Salas-Cobos E, Flores S, Patz JA: Linking deforestation to malaria in the Amazon: characterization of the breeding habitat of the principal malaria vector, Anopheles darlingi. Am J Trop Med Hyg 2009, 81:5-12. 78.Walsh JF, Molyneux DH, Birley MH: Deforestation: effects on vector-borne disease. Parasitology 1993, 106(Suppl):S55-S75. 79.Taylor D: Seeing the forests for the more than the trees. Environ Health Perspect 1997, 105:1186-1191. 80.Das NG, Talukdar PK, Das SC: Epidemiological and entomological aspects of malaria in forest-fringed villages of Sonitpur district, Assam. J Vector Borne Dis 2004, 41:5-9. 81.Elliott R: The influence of vector behavior on malaria transmission. Am J Trop Med Hyg 1972, 21:755-763. 82.Vittor AY, Gilman RH, Tielsch J, Glass G, Shields T, Lozano WS, Pinedo-Cancino V, Patz JA: The effect of deforestation on the human-biting rate of Anopheles darlingi, the primary vector of Falciparum malaria in the Peruvian Amazon. Am J Trop Med Hyg 2006, 74:3-11. 83.Smith DL, Ruktanonchai N: Progress in modelling malaria transmission. Adv Exp Med Biol 2010, 673:1-12. 84.Baimai V, Kijchalao U, Sawadwongporn P, Green CA: Geographic distribution and biting behaviour of four species of the Anopheles dirus complex (Diptera: Culicidae) in Thailand. Southeast Asian J Trop Med Public Health 1988, 19:151-161. 85.Dantur Juri MJ, Claps GL, Santana M, Zaidenberg M, Almiron WR: Abundance patterns of Anopheles pseudopunctipennis and Anopheles argyritarsis in northwestern Argentina. Acta Trop 2010, 115:234-241. 86.Hogg JC, Thomson MC, Hurd H: Comparative fecundity and associated factors for two sibling species of the Anopheles gambiae complex occurring sympatrically in The Gambia. Med Vet Entomol 1996, 10:385-391. 87.Sinka ME, Bangs MJ, Manguin S, Rubio-Palis Y, Chareonviriyaphap T, Coetzee M, Mbogo CM, Hemingway J, Patil AP, Temperley WH, Gething PW, Kabaria CW, Burkot TR, Harbach RE, Hay SI: A global map of dominant malaria vectors. Parasit Vectors 2012, 5:69. 88.Kirby MJ, Lindsay SW: Responses of adult mosquitoes of two sibling species, Anopheles arabiensis and A. gambiae s.s. (Diptera: Culicidae), to high temperatures. Bull Entomol Res 2004, 94:441-448. 89.Coetzee M, Craig M, le Sueur D: Distribution of African malaria mosquitoes belonging to the Anopheles gambiae complex. Parasitol Today 2000, 16:74-77. 90.Ebenezer A, Okiwelu SN, Agi PI, Noutcha MA, Awolola TS, Oduola AO: Species composition of the Anopheles gambiae complex across eco-vegetational zones in Bayelsa State, Niger Delta region, Nigeria. J Vector Borne Dis 2012, 49:164-167. 91.Singhasivanon P, Thimasarn K, Yimsamran S, Linthicum K, Nualchawee K, Dawreang D, Kongrod S, Premmanisakul N, Maneeboonyang W, Salazar N: Malaria in tree crop plantations in south-eastern and western provinces of Thailand. Southeast Asian J Trop Med Public Health 1999, 30:399-404. 92.Tanga MC, Ngundu WI, Tchouassi PD: Daily survival and human blood index of major malaria vectors associated with oil palm cultivation in Cameroon and their role in malaria transmission. Trop Med Int Health 2011, 16:447-457. 93.Pluess B, Mueller I, Levi D, King G, Smith TA, Lengeler C: Malaria–a major health problem within an oil palm plantation around Popondetta. Papua New Guinea Malar J 2009, 8:56. 94.Chang MS, Hii J, Buttner P, Mansoor F: Changes in abundance and behaviour of vector mosquitoes induced by land use during the development of an oil palm plantation in Sarawak. Trans R Soc Trop Med Hyg 1997, 91:382-386. 95.Bigoga JD, Nanfack FM, Awono-Ambene PH, Patchoke S, Atangana J, Otia VS, Fondjo E, Moyou RS, Leke RG: Seasonal prevalence of malaria vectors and entomological inoculation rates in the rubber cultivated area of Niete. South Region of Cameroon Parasit Vectors 2012, 5:197. 96.Basurko C, Demattei C, Han-Sze R, Grenier C, Joubert M, Nacher M, Carme B: Deforestation, agriculture and farm jobs: a good recipe for Plasmodium vivax in French Guiana. Malar J 2013, 12:90. 97.Wangroongsarb P, Sudathip P, Satimai W: Characteristics and malaria prevalence of migrant populations in malaria-endemic areas along the Thai-Cambodian border. Southeast Asian J Trop Med Public Health 2012, 43:261-269. 98.Sinka ME, Bangs MJ, Manguin S, Coetzee M, Mbogo CM, Hemingway J, Patil AP, Temperley WH, Gething PW, Kabaria CW, Burkot TR, Harbach RE, Hay SI: The dominant Anopheles vectors of human malaria in Africa, Europe and the Middle East: occurrence data, distribution maps and bionomic precis. Parasit Vectors 2010, 3:117. 99.Ngom EH, Ndione J-A, Ba Y, Konate L, Faye O, Diallo M, Dia I: Spatio-temporal analysis of host preferences and feeding patterns of malaria vectors in the sylvo-pastoral area of Senegal: impact of landscape classes. Parasit Vectors 2013, 6:332. 100.Massebo F, Balkew M, Gebre-Michael T, Lindtjorn B: Blood meal origins and insecticide susceptibility of Anopheles arabiensis from Chano in South-West Ethiopia. Parasit Vectors 2013, 6:44. 101.Subbarao SK, Sharma VP: Anopheline species complexes & malaria control. Indian J Med Res 1997, 106:164-173. 102.Rosenberg R, Maheswary NP: Forest malaria in Bangladesh. II Transmission by Anopheles dirus. Am J Trop Med Hyg 1982, 31:183-191. 103.Vatandoost H, Emami SN, Oshaghi MA, Abai MR, Raeisi A, Piazzak N, Mahmoodi M, Akbarzadeh K, Sartipi M: Ecology of malaria vector Anopheles culicifacies in a malarious area of Sistan va Baluchestan province, south-east Islamic Republic of Iran. E Mediterr Health J = La revue de sante de la Mediterranee orientale = al-Majallah al-sihhiyah li-sharq al-mutawassit 2011, 17:439-445. 104.Singh N, Singh OP, Sharma VP: Dynamics of malaria transmission in forested and deforested regions of Mandla District, central India (Madhya Pradesh). J Am Mosq Control Assoc 1996, 12:225-234. 105.Okwa OO, Akinmolayan FI, Carter V, Hurd H: Transmission dynamics of malaria in four selected ecological zones of Nigeria in the rainy season. Ann Afr Med 2009, 8:1-9. 106.Tirados I, Costantini C, Gibson G, Torr SJ: Blood-feeding behaviour of the malarial mosquito Anopheles arabiensis: implications for vector control. Med Vet Entomol 2006, 20:425-437. 107.Fontenille D, Lepers JP, Coluzzi M, Campbell GH, Rakotoarivony I, Coulanges P: Malaria transmission and vector biology on Sainte Marie Island, Madagascar. J Med Entomol 1992, 29:197-202. 108.Awolola TS, Okwa O, Hunt RH, Ogunrinade AF, Coetzee M: Dynamics of the malaria-vector populations in coastal Lagos, south-western Nigeria. Ann Trop Med Parasitol 2002, 96:75-82. 109.Afrane YA, Little TJ, Lawson BW, Githeko AK, Yan G: Deforestation and vectorial capacity of Anopheles gambiae Giles mosquitoes in malaria transmission, Kenya. Emerg Infect Dis 2008, 14:1533-1538. 110.Oyewole IO, Awolola TS, Ibidapo CA, Oduola AO, Okwa OO, Obansa JA: Behaviour and population dynamics of the major anopheline vectors in a malaria endemic area in southern Nigeria. J Vector Borne Dis 2007, 44:56-64. 111.Nanda N, Yadav RS, Subbarao SK, Joshi H, Sharma VP: Studies on Anopheles fluviatilis and Anopheles culicifacies sibling species in relation to malaria in forested hilly and deforested riverine ecosystems in northern Orissa, India. J Am Mosq Control Assoc 2000, 16:199-205. 112.Socheath S, Seng C, RathT S, Deesin V, Deesin T, Apiwathanasorn C: Study on bionomics of principal malaria vectors in Kratie province, Cambodia. Southeast Asian J Trop Med Public Health 2000, 31(Suppl 1):106-110. 113.Charlwood JD: Biological variation in Anopheles darlingi Root. Mem Inst Oswaldo Cruz 1996, 91:391-398. 114.Tadei WP, Dutary Thatcher B: Malaria vectors in the Brazilian amazon: Anopheles of the subgenus Nyssorhynchus. Rev Inst Med Trop Sao Paulo 2000, 42:87-94. 115.Baird JK: Resistance to therapies for infection by Plasmodium vivax. Clin Microbiol Rev 2009, 22:508-534. 116.Pampana E: A Textbook of Malaria Eradication. London: Oxford U.P.; 1969:1-593. 117.Oaks SC: Malaria: Obstacles and Opportunities: A Report of the Committee for the Study on Malaria Prevention and Control: Status Review and Alternative Strategies, Division of International Health, Institute of Medicine. Washington, D.C.: National Academy Press; 1991:1-309. 118.Lardeux F, Loayza P, Bouchite B, Chavez T: Host choice and human blood index of Anopheles pseudopunctipennis in a village of the Andean valleys of Bolivia. Malar J 2007, 6:8. 119.Robert V, Le Goff G, Toto JC, Essong J, Verhave JP: Early sporogonic development in local vectors of Plasmodium falciparum in rural Cameroon. Mem Inst Oswaldo Cruz 1994, 89 (Suppl 2):23-26. 120.Bellan SE: The importance of age dependent mortality and the extrinsic incubation period in models of mosquito-borne disease transmission and control. PLoS one 2010, 5:e10165. 121.Cox-Singh J, Davis TM, Lee KS, Shamsul SS, Matusop A, Ratnam S, Rahman HA, Conway DJ, Singh B: Plasmodium knowlesi malaria in humans is widely distributed and potentially life threatening. Clin Infect Dis 2008, 46:165-171. 122.Van der Palen M, Gillet P, Bottieau E, Cnops L, Van Esbroeck M, Jacobs J: Test characteristics of two rapid antigen detection tests (SD FK50 and SD FK60) for the diagnosis of malaria in returned travellers. Malar J 2009, 8:90. 123.Bharti PK, Chand SK, Singh MP, Mishra S, Shukla MM, Singh R, Singh N: Emergence of a new focus of Plasmodium malariae in forest villages of district Balaghat, Central India: implications for the diagnosis of malaria and its control. Trop Med Int Health 2013, 18:12-17. 124.Lee KS, Divis PC, Zakaria SK, Matusop A, Julin RA, Conway DJ, Cox-Singh J, Singh B: Plasmodium knowlesi: reservoir hosts and tracking the emergence in humans and macaques. PLoS Pathog 2011, 7:e1002015. 125.Kalra NL: Emergence of malaria zoonosis of simian origin as natural phenomenon in Greater Nicobars, Andaman & Nicobar islands–a preliminary note. J Commun Dis 1980, 12:6. 126.Deane LM, Deane MP, Ferreira Neto J: Studies on transmission of simian malaria and on a natural infection of man with Plasmodium simium in Brazil. Bull World Health Organ 1966, 35:805-808. 127.Deane LM: Simian malaria in Brazil. Mem Inst Oswaldo Cruz 1992, 87(Suppl 3):1-20. 128.da Silva-Nunes M, Moreno M, Conn JE, Gamboa D, Abeles S, Vinetz JM, Ferreira MU: Amazonian malaria: asymptomatic human reservoirs, diagnostic challenges, environmentally driven changes in mosquito vector populations, and the mandate for sustainable control strategies. Acta Trop 2012, 121:281-291. 129.Bousema T, Drakeley C: Epidemiology and infectivity of Plasmodium falciparum and Plasmodium vivax gametocytes in relation to malaria control and elimination. Clin Microbiol Rev 2011, 24:377-410. 130.Nkoghe D, Akue JP, Gonzalez JP, Leroy EM: Prevalence of Plasmodium falciparum infection in asymptomatic rural Gabonese populations. Malar J 2011, 10:33. 131.Drakeley CJ, Akim NI, Sauerwein RW, Greenwood BM, Targett GA: Estimates of the infectious reservoir of Plasmodium falciparum malaria in The Gambia and in Tanzania. Trans R Soc Trop Med Hyg 2000, 94:472-476. 132.Alves FP, Gil LH, Marrelli MT, Ribolla PE, Camargo EP, Da Silva LH: Asymptomatic carriers of Plasmodium spp. as infection source for malaria vector mosquitoes in the Brazilian Amazon. J Med Entomol 2005, 42:777-779. 133.Alves FP, Durlacher RR, Menezes MJ, Krieger H, Silva LH, Camargo EP: High prevalence of asymptomatic Plasmodium vivax and Plasmodium falciparum infections in native Amazonian populations. Am J Trop Med Hyg 2002, 66:641-648. 134.Babiker HA, Abdel-Muhsin AM, Ranford-Cartwright LC, Satti G, Walliker D: Characteristics of Plasmodium falciparum parasites that survive the lengthy dry season in eastern Sudan where malaria transmission is markedly seasonal. Am J Trop Med Hyg 1998, 59:582-590. 135.Bousema JT, Gouagna LC, Drakeley CJ, Meutstege AM, Okech BA, Akim IN, Beier JC, Githure JI, Sauerwein RW: Plasmodium falciparum gametocyte carriage in asymptomatic children in western Kenya. Malar J 2004, 3:18. 136.Pethleart A, Prajakwong S, Suwonkerd W, Corthong B, Webber R, Curtis C: Infectious reservoir of Plasmodium infection in Mae Hong Son Province, north-west Thailand. Malar J 2004, 3:34. 137.Wells TN, Burrows JN, Baird JK: Targeting the hypnozoite reservoir of Plasmodium vivax: the hidden obstacle to malaria elimination. Trends Parasitol 2010, 26:145-151. 138.Das SC, Baruah I: Incrimination of Anopheles minimus Theobald and Anopheles balabacensis balabacensis Baisas (A. dirus) as malaria vectors in Mizoram. Indian J Malariol 1985, 22:53-55. 139.Adak DK: Demography and Health Profile of the Tribals : A Study of M.P. New Delhi: Anmol Publications; 2003. 140.Peeters Grietens K, Xuan XN, Van Bortel W, Duc TN, Ribera JM, Ba Nhat T, Van KP, Le Xuan H, D’Alessandro U, Erhart A: Low perception of malaria risk among the Ra-glai ethnic minority in south-central Vietnam: implications for forest malaria control. Malar J 2010, 9:23. 141.Myers N: Tropical forests: present status and future outlook. Clim Change 1991, 19:3-32. 142.Yadav SP, Kalundha RK, Sharma RC: Sociocultural factors and malaria in the desert part of Rajasthan, India. J Vector Borne Dis 2007, 44:205-212. 143.Hossain SM, Bhuiya A, Rasheed S: Correlates of perceived malarial episodes and treatment-seeking behavior in a malaria-endemic rural area in Bangladesh. Southeast Asian J Trop Med Public Health 2001, 32:707-719. 144.Lefevre T, Gouagna LC, Dabire KR, Elguero E, Fontenille D, Renaud F, Costantini C, Thomas F: Beer consumption increases human attractiveness to malaria mosquitoes. PLoS one 2010, 5:e9546. 145.Somboon P, Aramrattana A, Lines J, Webber R: Entomological and epidemiological investigations of malaria transmission in relation to population movements in forest areas of north-west Thailand. Southeast Asian J Trop Med Public Health 1998, 29:3-9. 146.Martens P, Hall L: Malaria on the move: human population movement and malaria transmission. Emerg Infect Dis 2000, 6:103-109. 147.Sahu SS, Gunasekaran K, Jambulingam P: Bionomics of Anopheles minimus and An. fluviatilis (Diptera: Culicidae) in east-central India, endemic for falciparum malaria: human landing rates, host feeding, and parity. J Med Entomol 2009, 46:1045-1051. 148.Cai X, Deng D, Wu K, Tang L, Lan C, Gu Z, He Y, Wang K, Wu D, Du J: A study on human behavior and socioeconomic factors affecting malaria transmission and control in Qiongzhong, Hainan. Zhongguo ji sheng chong xue yu ji sheng chong bing za zhi = Chin J Parasitol Parasit Dis 1995, 13:89-93. 149.Butraporn P, Sornmani S, Hungsapruek T: Social, behavioural, housing factors and their interactive effects associated with malaria occurrence in east Thailand. Southeast Asian J Trop Med Public Health 1986, 17:386-392. 150.Radford AJ, Van Leeuwen H, Christian SH: Social aspects in the changing epidemiology of malaria in the highlands of New Guinea. Ann Trop Med Parasitol 1976, 70:11-23. 151.Sanh NH, Van Dung N, Thanh NX, Trung TN, Van Co T, Cooper RD: Forest malaria in central Vietnam. Am J Trop Med Hyg 2008, 79:652-654. 152.Das NG, Bhuyan M, Das SC: Some observations on malaria in tribal areas of Bastar District, Chhattisgarh. J Commun Dis 2003, 35:300-305. 153.Kosinski LA: Governments’ perceptions and policies of population redistribution in East and South East Asia. Popul Geogr 1981, 3:4-15. 154.Lindsay SW, Martens WJ: Malaria in the African highlands: past, present and future. Bull World Health Organ 1998, 76:33-45. 155.Dahesh SM, Bassiouny HK, El-Masry SA: Socioeconomic and environmental factors affecting malaria infection in Fayoum Governorate, Egypt. J Egypt Soc Parasitol 2009, 39:511-523. 156.Ghebreyesus TA, Haile M, Witten KH, Getachew A, Yohannes M, Lindsay SW, Byass P: Household risk factors for malaria among children in the Ethiopian highlands. Trans R Soc Trop Med Hyg 2000, 94:17-21. 157.Garros C, Van Bortel W, Trung HD, Coosemans M, Manguin S: Review of the Minimus Complex of Anopheles, main malaria vector in Southeast Asia: from taxonomic issues to vector control strategies. Trop Med Int Health 2006, 11:102-114. 158.Singh N, Dash AP, Thimasarn K: Fighting malaria in Madhya Pradesh (Central India): are we losing the battle? Malar J 2009, 8:93. 159.Giglioli G: Ecological change as a factor in renewed malaria transmission in an eradicated area. A localized outbreak of A. aquasalis transmitted malaria on the demerara river estuary, British Guiana, in the fifteenth year of A. darlingi and malaria eradication. Bull World Health Organ 1963, 29:131-145. 160.Dossou-Yovo J, Ouattara A, Doannio JM, Diarrassouba S, Chauvancy G: Malaria surveys in a humid savannah region in Cote d’Ivoire. Med Trop (Mars) 1998, 58:51-56. 161.Feyisetan BJ, Asa S, Ebigbola JA: Mothers’ management of childhood diseases in Yorubaland: the influence of cultural beliefs. Health Transit Rev 1997, 7:221-234. 162.Herndon CN, Uiterloo M, Uremaru A, Plotkin MJ, Emanuels-Smith G, Jitan J: Disease concepts and treatment by tribal healers of an Amazonian forest culture. J Ethnobiol Ethnomed 2009, 5:27. 163.Sharma SK, Jalees S, Kumar K, Rahman SJ: Knowledge, attitude and beliefs about malaria in a tribal area of Bastar district (Madhya Pradesh). Indian J Public Health 1993, 37:129-132. 164.Manguin S: Biodiversity of Malaria in the World. Montrouge: John Libbey Eurotext; 2008. 165.Adera TD: Beliefs and traditional treatment of malaria in Kishe settlement area, southwest Ethiopia. Ethiop Med J 2003, 41:25-34. 166.Alves MJ, Barata LC, Barata Rde C, de Almeida Mdo C, Gutierrez EB, Wanderley DM, de Andrade JC: Socioeconomic aspects of subjects with imported malaria in the metropolitan area of Sao Paulo, Brazil. I. Characterization of the population and knowledge about the disease. Rev Saude Publica 1990, 24:253-258. 167.Dhillon HS, Kar SB: Malaria eradication and investigation of cultural patterns and beliefs among tribal population in india. Int J Health Educ 1965, Ill:31-40. 168.Deressa W, Ali A, Hailemariam D: Malaria-related health-seeking behaviour and challenges for care providers in rural Ethiopia: implications for control. J Biosoc Sci 2008, 40:115-135. 169.Elphick H, Elphick D: Factors that contribute to the low use of bed nets in a malaria endemic zone of sub-Saharan Africa: a questionnaire survey in a rural population in Zambia. Cent Afr J Med 2003, 49:87-89. 170.Pulford J, Hetzel MW, Bryant M, Siba PM, Mueller I: Reported reasons for not using a mosquito net when one is available: a review of the published literature. Malar J 2011, 10:83. 171.Esse C, Utzinger J, Tschannen AB, Raso G, Pfeiffer C, Granado S, Koudou BG, N’Goran EK, Cisse G, Girardin O, Tanner M, Obrist B: Social and cultural aspects of ‘malaria’ and its control in central Cote d’Ivoire. Malar J 2008, 7:224. 172.Moore SJ, Min X, Hill N, Jones C, Zaixing Z, Cameron MM: Border malaria in China: knowledge and use of personal protection by minority populations and implications for malaria control: a questionnaire-based survey. BMC Public Health 2008, 8:344. 173.Gosling R, Drakeley C, Mwita A, Chandramohan D: Presumptive treatment of fever cases as malaria: help or hindrance for malaria control? Malar J 2008, 7:132. 174.Talisuna AO, Langi P, Bakyaita N, Egwang T, Mutabingwa TK, Watkins W, Van Marck E, D’Alessandro U: Intensity of malaria transmission, antimalarial-drug use and resistance in Uganda: what is the relationship between these three factors? Trans R Soc Trop Med Hyg 2002, 96:310-317. 175.Hastings IM, Watkins WM: Intensity of malaria transmission and the evolution of drug resistance. Acta Trop 2005, 94:218-229. 176.Alam MT, Vinayak S, Congpuong K, Wongsrichanalai C, Satimai W, Slutsker L, Escalante AA, Barnwell JW, Udhayakumar V: Tracking origins and spread of sulfadoxine-resistant Plasmodium falciparum dhps alleles in Thailand. Antimicrob Agents Chemother 2011, 55:155-164.
|